Terminal deoxynucleotidyl transferase (TdT), also known as DNA nucleotidylexotransferase (DNTT) or terminal transferase, is a specialized DNA polymerase expressed in immature, pre-B, pre-T lymphoid cells, and acute lymphoblastic leukemia/lymphoma cells. TdT adds N-nucleotides to the V, D, and Jexons of the TCR and BCR genes during antibody gene recombination, enabling the phenomenon of junctional diversity. In humans, terminal transferase is encoded by the DNTTgene.[5][6] As a member of the X family of DNA polymerase enzymes, it works in conjunction with polymerase λ and polymerase μ, both of which belong to the same X family of polymerase enzymes. The diversity introduced by TdT has played an important role in the evolution of the vertebrate immune system, significantly increasing the variety of antigen receptors that a cell is equipped with to fight pathogens. Studies using TdT knockout mice have found drastic reductions (10-fold) in T-cell receptor (TCR) diversity compared with that of normal, or wild-type, systems. The greater diversity of TCRs that an organism is equipped with leads to greater resistance to infection.[7][8] Although TdT was one of the first DNA polymerases identified in mammals in 1960,[9] it remains one of the least understood of all DNA polymerases.[7] In 2016–18, TdT was discovered to demonstrate in trans template dependant behaviour in addition to its more broadly known template independent behaviour[10][11]
TdT is absent in fetal liver HSCs, significantly impairing junctional diversity in B-cells during the fetal period.[12]
Function and regulation
Generally, TdT catalyses the addition of nucleotides to the 3' terminus of a DNA molecule. Unlike most DNA polymerases, it does not require a template. The preferred substrate of this enzyme is a 3'-overhang, but it can also add nucleotides to blunt or recessed 3' ends. Further, TdT is the only polymerase that is known to catalyze the synthesis of 2-15nt DNA polymers from free nucleotides in solution in vivo.[13]In vitro, this behaviour catalyzes the general formation of DNA polymers without specific length.[14] The 2-15nt DNA fragments produced in vivo are hypothesized to act in signaling pathways related to DNA repair and/or recombination machinery.[13] Like many polymerases, TdT requires a divalent cation cofactor,[15] however, TdT is unique in its ability to use a broader range of cations such as Mg2+, Mn2+, Zn2+ and Co2+.[15] The rate of enzymatic activity depends on the available divalent cations and the nucleotide being added.[16]
TdT is expressed mostly in the primary lymphoid organs, like the thymus and bone marrow. Regulation of its expression occurs via multiple pathways. These include protein-protein interactions, like those with TdIF1. TdIF1 is another protein that interacts with TdT to inhibit its function by masking the DNA binding region of the TdT polymerase. The regulation of TdT expression also exists at the transcriptional level, with regulation influenced by stage-specific factors, and occurs in a developmentally restrictive manner.[7][17][18] Although expression is typically found to be in the primary lymphoid organs, recent work has suggested that stimulation via antigen can result in secondary TdT expression along with other enzymes needed for gene rearrangement outside of the thymus for T-cells.[19] Patients with acute lymphoblastic leukemia greatly over-produce TdT.[16] Cell lines derived from these patients served as one of the first sources of pure TdT and lead to the discovery that differences in activity exist between human and bovine isoforms.[16]
Mechanism
Graphic describing the mechanism of nucleotidyl condensation with ssDNA as catalyzed by terminal deoxynucleotidyl transferase with divalent cation cofactors. Two aspartate residues facilitate cation binding and nucleophilic attack.
Similar to many polymerases, the catalytic site of TdT has two divalent cations in its palm domain that assist in nucleotide binding, help lower the pKa of the 3'-OH group and ultimately facilitate the departure of the resultant pyrophosphate by-product.[20][21]
Isoform Variation
Several isoforms of TdT have been observed in mice, bovines, and humans. To date, two variants have been identified in mice while three have been identified in humans.[22]
The two splice variants identified in mice are named according to their respective lengths: TdTS consists of 509 amino acids while TdTL, the longer variant, consists of 529 amino acids. The differences between TdTS and TdTL occur outside regions that bind DNA and nucleotides. That the 20 amino acid difference affects enzymatic activity is controversial, with some arguing that TdTL's modifications bestow exonuclease activity while others argue that TdTL and TdTS have nearly identical in vitro activity. Additionally, TdTL reportedly can modulate the catalytic activity of TdTS in vivo through an unknown mechanism. It is suggested that this aids in the regulation of TdT's role in V(D)J recombination.[23]
Human TdT isoforms have three variants TdTL1, TdTL2, and TdTS. TdTL1 is broadly expressed in lymphoid cell lines while TdTL2 is predominantly expressed in normal small lymphocytes. Both localize in the nucleus when expressed[24] and both possess 3'->5' exonuclease activity.[25] In contrast, TdTS isoforms do not possess exonuclease activity and perform the necessary elongation during V(D)J recombination.[25] Since a similar exonuclease activity hypothesized in murine TdTL is found in human and bovine TdTL, some postulate that bovine and human TdTL isoforms regulate TdTS isoforms in a similar manner as proposed in mice.[23] Further, some hypothesize that TdTL1 may be involved in the regulation of TdTL2 and/or TdTS activity.
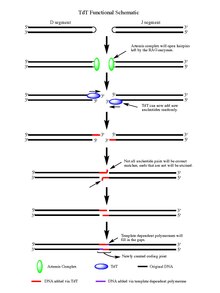
Role in V(D)J Recombination
This image provides a visual representation of how TdT works in the process of antibody gene rearrangement. Know that although the image uses D and J segments, the same type of rearrangements happen with other segment pairs as well.
Upon the action of the RAG 1/2 enzymes, the cleaved double-stranded DNA is left with hairpin structures at the end of each DNA segment created by the cleavage event. The hairpins are both opened by the Artemis complex, which has endonuclease activity when phosphorylated, providing the free 3' OH ends for TdT to act upon. Once the Artemis complex has done its job and added palindromic nucleotides (P-nucleotides) to the newly opened DNA hairpins, the stage is set for TdT to do its job. TdT is now able to come in and add N-nucleotides to the existing P-nucleotides in a 5' to 3' direction that polymerases are known to function. On average 2-5 random base pairs are added to each 3' end generated after the action of the Artemis complex. The number of bases added is enough for the two newly synthesized ssDNA segments to undergo microhomology alignment during non-homologous end joining according to the normal Watson-Crick base pairing patterns (A-T, C-G). From there unpaired nucleotides are excised by an exonuclease, like the Artemis Complex (which has exonuclease activity in addition to endonuclease activity), and then template-dependent polymerases can fill the gaps, finally creating the new coding joint with the action of ligase to combine the segments. Although TdT does not discriminate among the four base pairs when adding them to the N-nucleotide segments, it has shown a bias for guanine and cytosine base pairs.[7]
Template Dependent Activity
TDT bound to three DNA strands demonstrating the active configuration of its template dependant catalysis.
In a template-dependant manner, TdT can incorporate nucleotides across strand breaks in double-stranded DNA in a manner referred to as in trans in contrast to the in cis mechanism found in most polymerases. This occurs optimally with a one base-pair break between strands and less so with an increasing gap. This is facilitated by a subsection of TdT called Loop1 which selectively probes for short breaks in double-stranded DNA. Further, the discovery of this template dependant activity has led to more convincing mechanistic hypotheses as to how the distribution of lengths of the additions of the N regions arise in V(D)J recombination.[26]
A graphical diagram depicting the in trans template dependant activity of terminal deoxynucleotidyl transferase. Loop1 is highlighted in red.
Polymerase μ and polymerase λ exhibit similar in trans templated dependant synthetic activity to TdT, but without similar dependence on downstream double-stranded DNA.[27] Further, Polymerase λ has also been found to exhibit similar template-independent synthetic activity. Along with activity as a terminal transferase, it is known to also work in a more general template-dependent fashion.[28] The similarities between TdT and polymerase μ suggest they are closely evolutionarily related.[26]
Uses
Terminal transferase has applications in molecular biology. It can be used in RACE to add nucleotides that can then be used as a template for a primer in subsequent PCR. It can also be used to add nucleotides labeled with radioactiveisotopes, for example in the TUNEL assay (Terminal deoxynucleotidyl transferase dUTP Nick End Labeling) for the demonstration of apoptosis (which is marked, in part, by fragmented DNA). It is also used in the immunofluorescence assay for the diagnosis of acute lymphoblastic leukemia.[29]
In immunohistochemistry and flow cytometry, antibodies to TdT can be used to demonstrate the presence of immature T and B cells and pluripotent hematopoietic stem cells, which possess the antigen, while mature lymphoid cells are always TdT-negative. While TdT-positive cells are found in small numbers in healthy lymph nodes and tonsils, the malignant cells of acute lymphoblastic leukemia are also TdT-positive, and the antibody can, therefore, be used as part of a panel to diagnose this disease and to distinguish it from, for example, small cell tumors of childhood.[30]
TdT has also seen recent application in the De Novo synthesis of oligonucleotides, with TdT-dNTP tethered analogs capable of primer extension by 1 nt at a time.[31] In other words, the enzyme TdT has demonstrated the capability of making synthetic DNA by adding one letter at a time to a primer sequence.
^Yang-Feng TL, Landau NR, Baltimore D, Francke U (1986). "The terminal deoxynucleotidyltransferase gene is located on human chromosome 10 (10q23----q24) and on mouse chromosome 19". Cytogenetics and Cell Genetics. 43 (3–4): 121–6. doi:10.1159/000132309. PMID3467897.
^Hardy R (2008). "Chapter 7: B Lymphocyte Development and Biology". In Paul W (ed.). Fundamental Immunology (Book) (6th ed.). Philadelphia: Lippincott Williams & Wilkins. pp. 237–269. ISBN978-0-7817-6519-0.
^ abRamadan K, Shevelev IV, Maga G, Hübscher U (May 2004). "De novo DNA synthesis by human DNA polymerase lambda, DNA polymerase mu and terminal deoxyribonucleotidyl transferase". Journal of Molecular Biology. 339 (2): 395–404. doi:10.1016/j.jmb.2004.03.056. PMID15136041.
^Bollum FJ, Chang LM, Tsiapalis CM, Dorson JW (1974). "[8] Nucleotide polymerizing enzymes from calf thymus gland". Nucleotide polymerizing enzymes from calf thymus gland. Methods in Enzymology. Vol. 29. pp. 70–81. doi:10.1016/0076-6879(74)29010-4. ISBN9780121818920. PMID4853390.
^Cherrier M, D'Andon MF, Rougeon F, Doyen N (February 2008). "Identification of a new cis-regulatory element of the terminal deoxynucleotidyl transferase gene in the 5' region of the murine locus". Molecular Immunology. 45 (4): 1009–17. doi:10.1016/j.molimm.2007.07.027. PMID17854898.
^Zhang Y, Shi M, Wen Q, Luo W, Yang Z, Zhou M, et al. (2012-01-01). "Antigenic stimulation induces recombination activating gene 1 and terminal deoxynucleotidyl transferase expression in a murine T-cell hybridoma". Cellular Immunology. 274 (1–2): 19–25. doi:10.1016/j.cellimm.2012.02.008. PMID22464913.
^Steenberg ML, Lokhandwala MF, Jandhyala BS (1988). "Abnormalities in the sodium transport as the causative factor for enhanced norepinephrine overflow in the spontaneously hypertensive rat". Clinical & Experimental Hypertension, Part A. 10 (5): 833–41. doi:10.1080/07300077.1988.11878788. PMID2846215.
^ abBland RD, Clarke TL, Harden LB (February 1976). "Rapid infusion of sodium bicarbonate and albumin into high-risk premature infants soon after birth: a controlled, prospective trial". American Journal of Obstetrics and Gynecology. 124 (3): 263–7. doi:10.1016/0002-9378(76)90154-x. PMID2013.
O'Malley DP, Orazi A (August 2006). "Terminal deoxynucleotidyl transferase-positive cells in spleen, appendix and branchial cleft cysts in pediatric patients". Haematologica. 91 (8): 1139–40. PMID16885057.
Yamashita N, Shimazaki N, Ibe S, Kaneko R, Tanabe A, Toyomoto T, et al. (July 2001). "Terminal deoxynucleotidyltransferase directly interacts with a novel nuclear protein that is homologous to p65". Genes to Cells. 6 (7): 641–52. doi:10.1046/j.1365-2443.2001.00449.x. PMID11473582. S2CID19573920.{{cite journal}}: CS1 maint: overridden setting (link)
Chang LM, Bollum FJ (1986). "Molecular biology of terminal transferase". CRC Critical Reviews in Biochemistry. 21 (1): 27–52. doi:10.3109/10409238609113608. PMID3524991.
Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (October 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID9373149.
Shimazaki N, Fujita K, Koiwai O (March 2002). "[Expression and function of terminal deoxynucleotidyl-transferase and discovery of novel DNA polymerase mu]". Seikagaku. The Journal of Japanese Biochemical Society. 74 (3): 227–32. PMID11974916.