馬許在他1892年的論文描繪了角鼻龍首個骨架重建圖,身長6.7公尺、身高3.7公尺。[5]吉爾摩在1920年指出,前者的重建把軀幹弄得太長,至少多加了六節脊椎。該錯誤在隨後多篇文獻中反覆提及,包括1899年(1920年才出版)邦德(Frank Bond)在奈特(英语:Charles R. Knight)指導下繪製的首件生前復原圖。1901年出版更準確的復原圖是由格里森(英语:Joseph M. Gleeson)同樣於奈特監督下製作。吉爾摩於1910至1911年間裝設了正模骨架,並展示於國立自然史博館。多數早期重建都把角鼻龍描繪成尾巴拖地的直立站姿。[2]:115–116相比之下吉爾摩的重建在當時頗為前衛:[6]:276由大腿骨上端得到啟發,發現其與小腿成一定角度,而形成水平而非直立、尾部懸空、能夠奔跑的姿勢。因為化石受壓縮變形的程度嚴重,吉爾摩難以架設立體骨骼,而用淺浮雕的樣貌展示於牆面上。[2]:114部分骨骼嵌入牆面,使要進行更深入研究受到阻礙。在2014至2019年博物館恐龍展示翻修期間,館方將標本從牆面取下,[7][8]並於2019年計畫開幕的新展覽中,以一個新製作的立體複製品取而代之;原始標本則回歸博物館館藏,使學者得以徹底研究。[8]
北美其他發現
自模式種的正模發現以後,直到1960年代才再次發現另一具較重要的角鼻龍化石,是由詹姆斯·麥德森(英语:James Henry Madsen)及團隊在猶他克里夫蘭羅伊採石場挖出破碎、關節脫落的部分骨骼及頭骨UMNH VP 5278標本,是體型最大的角鼻龍標本之一。[9]:211976年艾瑞克森(Thor Erikson)在科羅拉多果城發現第二個關節連接、含頭骨的標本MWC 1。[10]這是一隻相對完整的骨骼,缺乏下頜、手臂及腹肋。雖然頭骨較完整,發現時已呈關節脫落狀態,側面壓得很扁。來自一隻大型個體,但由頭骨縫線開放程度顯示,其尚未達到成熟的體型。[9]:2–3提供博物館展示的科學準確3D頭骨重建模型,製作過程很複雜,從單一件原始骨骼翻模、矯正變形部位、重建遺失部分、將骨骼模型安裝到適當位置、最後上色以還原骨骼原始色彩。[11]
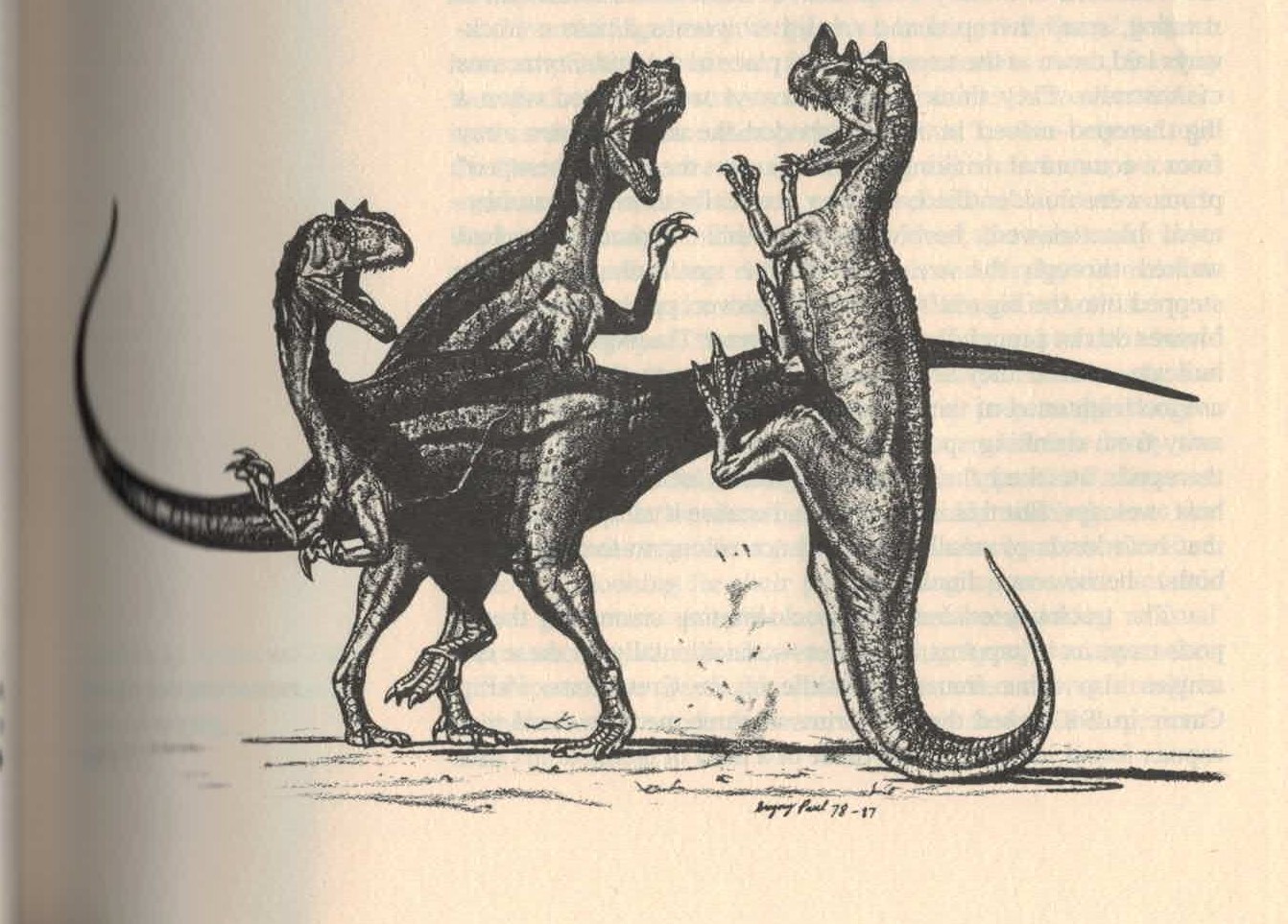
1884年馬許認為角鼻龍的鼻角是作為攻擊與防禦的強大武器,1920年吉爾摩認同其論點。[3]:331[2]:82然而現代普遍認為鼻角不太可能當作武器。[10]1985年大衛·諾曼(英语:David B. Norman)認為鼻角可能並非用來抵禦其他掠食者,而是爭奪配偶的雄性角鼻龍種內打鬥使用。[54]1988年奎格里·保羅(英语:Gregory S. Paul)提出類似功能,並繪製兩隻角鼻龍進行一場非致命的互撞競賽。[6]1990年羅威和高席爾提出更進一步的論點,認為鼻角僅具有視覺展示功能,完全無法打鬥。[19]為了達到炫耀目的,鼻角可能具有鮮豔色彩。[31]另外沿身體中線生長的成排皮甲也可能有展示功能。[19]
葡萄牙洛里尼揚層新港段(Porto Novo Member)發現角鼻龍的破碎遺骸透露了該物種也許存在於此地。洛里尼揚層的恐龍生態組成結構與莫里遜層幾乎一樣,或至少都是類似物種。[62]除了角鼻龍,葡萄牙岩層也同樣出土過來自北美的異特龍和蠻龍,至於洛里尼揚諾龍只在葡萄牙發現過。新港段的植食性恐龍包括蜥腳類的迪涅魯龍、洛里尼揚龍、茲必龍;劍龍類的米拉加亞龍。[66][24][25]在晚侏羅世,歐洲與北美之間僅隔著剛形成的狹窄大西洋,而葡萄牙所在的伊比利半島當時是一座大型孤島,與歐洲其他地區隔離開來。根據馬諦尤斯等人所述,葡萄牙與北美獸腳類種群的相似程度,代表之間曾經存在過陸橋使動物得以互相遷徙。[24][25]不過馬拉法雅等人提出不同、更為複雜的情境假設,因為葡萄牙其他動物如蜥腳類、海龜、鱷類都與北美有明顯差異,代表當陸塊開始分開時,某些物種是透過交換遷徙來的,某些則在當地產生特有化。[26]
1920年代以後,角鼻龍形象長期固化,後續的畫家皆參考格里森或斯密,逐漸完全脫離動物的真實形象。直到1980年代突然起了變革,奎格里·保羅(英语:Gregory S. Paul)觀察了正模標本,將角鼻龍詮釋為行動活躍的水平姿態,更貼近現代形象。其中一幅 (页面存档备份,存于互联网档案馆)角鼻龍以尾巴撐地抬起雙腳踢擊兩隻異特龍,類似袋鼠的行為。另一幅 (页面存档备份,存于互联网档案馆)依照他對鼻角功能的論點,展現了角鼻龍透過鼻角側擊對抗同類。這些成為新式古生物形象的代表性作品,並在1990年代佔據主流地位。[6]
^Tidwell, V. Restoring crushed Jurassic dinosaur skulls for display. The Continental Jurassic. Museum of Northern Arizona Bulletin. 1996, 60.
^Britt, B.B.; Chure, D.J.; Holtz, T.R., Jr.; Miles, C.A.; Stadtman, K.L. A reanalysis of the phylogenetic affinities of Ceratosaurus (Theropoda, Dinosauria) based on new specimens from Utah, Colorado, and Wyoming. Journal of Vertebrate Paleontology. 2000, 20 (suppl): 32A. doi:10.1080/02724634.2000.10010765.
^Rauhut, O.W.M. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology. 2003: 25.
^ 15.015.1Britt, B.B.; Miles, C.A.; Cloward, K.C.; Madsen, J.H. A juvenile Ceratosaurus (Theropoda, Dinosauria) from Bone Cabin Quarry West (Upper Jurassic, Morrison Formation), Wyoming. Journal of Vertebrate Paleontology. 1999, 19 (Supplement to No 3): 33A. doi:10.1080/02724634.1999.10011202.
^Foster, J. Appendix. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 2007: 327–329. ISBN 978-0-253-34870-8.
^ 17.017.1Zils, C.W.; Moritz, A.; Saanane, C. Tendaguru, the most famous dinosaur locality of Africa. Review, survey and future prospects. Documenta Naturae. 1995, 97: 1–41.
^ 18.018.118.218.3Janensch, W. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica. 1925,. Supplement VIII: 1–100 (德语).
^ 19.019.119.219.3Rowe, T.; Gauthier, J. Ceratosauria. Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). The Dinosauria. University of California Press. 1990: 151–168. ISBN 978-0-520-06726-4.
^ 21.021.121.2Rauhut, O.W.M. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Special Papers in Palaeontology. 2011, 86: 195–239.
^Huene, F.v. On several known and unknown reptiles of the order Saurischia from England and France. Annals and Magazine of Natural History. 1926,. Serie 9 (17): 473–489. doi:10.1080/00222932608633437.
^ 23.023.1Rauhut, O.W.M.; Milner, A.C.; Moore-Fay, S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zoological Journal of the Linnean Society. 2010, 158 (1): 155–195. doi:10.1111/j.1096-3642.2009.00591.x.
^ 24.024.124.2Mateus, O.; Antunes, M.T. Ceratosaurus (Dinosauria: Theropoda) in the Late Jurassic of Portugal. 31st International Geological Congress, Abstract Volume (Rio de Janeiro). 2000.
^ 25.025.125.225.3Mateus, O.; Walen, A.; Antunes, M.T. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science, Bulletin 36. 2006.
^ 26.026.126.2Malafaia, E.; Ortega, F.; Escaso, F.; Silva, B. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal. Historical Biology. October 3, 2015, 27 (7): 938–946. ISSN 0891-2963. doi:10.1080/08912963.2014.915820.
^Soto, M.; Perea, D. A ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic–Early Cretaceous of Uruguay. Journal of Vertebrate Paleontology. 2008, 28 (2): 439–444. doi:10.1671/0272-4634(2008)28[439:acdtft]2.0.co;2.
^Therrien, F.; Henderson, D.M. My theropod is bigger than yours ... or not: estimating body size from skull length in theropods. Journal of Vertebrate Paleontology. 2007, 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:mtibty]2.0.co;2.
^ 31.031.131.231.3Foster, J. Gargantuan to Minuscule: The Morrison Menagerie, Part II. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 2007: 162–242. ISBN 978-0-253-34870-8.
^ 32.032.132.232.3Henderson, D.M. Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods. Gaia. 1998, (15): 219–226.
^ 34.034.1Rauhut, O.W.M. Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia. Journal of Vertebrate Paleontology. 2004, 24 (4): 894–902. doi:10.1671/0272-4634(2004)024[0894:paaogs]2.0.co;2.
^ 35.035.1Carrano, M.T.; Choiniere, J. New information on the forearm and manus of Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda), with implications for theropod forelimb evolution. Journal of Vertebrate Paleontology. 2016, 36 (2): –1054497. doi:10.1080/02724634.2015.1054497.
^Hendrickx, C.; Hartman, S.A.; Mateus, O. An overview of non-avian theropod discoveries and classification. PalArch's Journal of Vertebrate Palaeontology. 2015, 12 (1): 1–73.
^ 44.044.1Foster, J.R. Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.. New Mexico Museum of Natural History and Science Bulletin 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2003: 29.
^ 46.046.1Foster, J.R.; Chure, D.J. Hindlimb allometry in the Late Jurassic theropod dinosaur Allosaurus, with comments on its abundance and distribution. New Mexico Museum of Natural History and Science Bulletin. 2006, 36: 119–122.
^Carpenter, Kenneth. Variation in a population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA. Paleontological Research. 2010, 14 (4): 250–259. doi:10.2517/1342-8144-14.4.250.
^Carpenter, Kenneth; Paul, Gregory S. Comment on Allosaurus Marsh, 1877 (Dinosauria, Theropoda): proposed conservation of usage by designation of a neotype for its type species Allosaurus fragilis Marsh, 1877. Bulletin of Zoological Nomenclature. 2015, 72: 1.
^Holliday, C.M.; Witmer, L.M. Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids. Journal of Vertebrate Paleontology. December 12, 2008, 28 (4): 1073–1088. doi:10.1671/0272-4634-28.4.1073.
^Sanders, R.K.; Smith, D.K. The endocranium of the theropod dinosaur Ceratosaurus studied with computer tomography. Acta Palaeontologica Polonica. 2005, 50 (3).
^ 57.057.157.2Molnar, R.E. Theropod paleopathology: a literature survey. Tanke, D.H.; Carpenter, K. (编). Mesozoic Vertebrate Life. Indiana University Press. 2001: 337–363.
^Rothschild, B.; Tanke, D.H.; Ford, T.L. Theropod stress fractures and tendon avulsions as a clue to activity. Tanke, D.H.; Carpenter, K. (编). Mesozoic Vertebrate Life. Indiana University Press. 2001: 331–336.
^Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations. Geological Society of America Abstracts with Programs. 2006, 38 (6): 7.
^Bilbey, S.A. Cleveland-Lloyd Dinosaur Quarry – age, stratigraphy and depositional environments. Carpenter, K.; Chure, D.; Kirkland, J.I. (编). The Morrison Formation: An Interdisciplinary Study. Modern Geology 22. Taylor and Francis Group. 1998: 87–120. ISSN 0026-7775.
^Russell, D.A. An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. 1989: 64–70. ISBN 978-1-55971-038-1.
^ 62.062.1Mateus, O. Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 223–231.
^Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
^Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. The fauna and flora of the Morrison Formation: 2006. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 233–248.
^Carpenter, K. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 131–138.
^Mateus, O.; Mannion, P.D.; Upchurch, P. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology. 2014, 34 (3): 618–634. doi:10.1080/02724634.2013.822875.
^Lucas, F. A. (1901). Animals of the Past. McClure, Phillips.
^Hutchinson, H. N. (1910). Extinct Monsters and Creatures of Other Days: A Popular Account of Some of the Larger Forms of Ancient Animal Life. Chapman & Hall.







_-_AMNH_-_DSC06292.JPG)











.jpg)



.jpg)













{kind=link}
{kind=link}
{kind=link}
{kind=link}